Chimpanzee mutation rate is largely determined by fathers' age and, overall, implies a Pan-Homo divergence rate of ~13 million years (95% CI: 11-17 Ma), about double than usually assumed by conservative scholastic inertia.
Oliver Venn et al., Strong male bias drives germline mutation in chimpanzees. Science 2014. Pay per view → LINK [doi:10.1126/science.344.6189.1272]
 |
| cc Matthew Hoelscher |
The focus of this study are the important differences between patrilineal and matrilineal mutation rate depending on the father's age among chimpanzees, notably more biased than among humans. However the resulting estimate for Pan-Homo divergence is not less important because it radically challenges the usual assumptions of 5-7 Ma, repeated once and again in molecular clock estimates, which are based on studies that are already quite obsolete.
In the studied captive population of Western chimpanzees 30 out of 35 mutations happened in the paternal lineage, and these increase with the father's age. No effect could be attributed to maternal age or familiar peculiarities.
Interestingly most of these patrilineal mutations happen near the telomeres, an effect not seen in female line mutations.
Owing to this gender bias, the mutation rate of the X chromosome among chimpanzees is 74% that of autosomal DNA (in humans: 85%).
The gender bias in mutation rate and its differential with humans is attributed to differences in mating systems among great apes, with chimpanzees having the greatest competition among males, what is reflected in testicle size. They predict that gorillas (who experience less competition between males) will show less patrilineal mutation rate bias than humans and chimpanzees.
This is probably the more synthetic paragraph from the study:
Under a model in which the mutation rate increases linearly with parental age, the rate of neutral substitution is the ratio of the average number of mutations inherited per generation to the average parental age. We predict the neutral substitution rate to be ~0.46 × 10−9 per base pair (bp) per year in chimpanzees, compared to estimates in humans of ~0.51 × 10−9 bp−1 year−1 (9). These results are consistent with near-identical levels of lineage-specific sequence divergence (12) but surprising given the differences in paternal age effect. In the intersection of the autosomal genome accessible in this study and regions where human and chimpanzee genomes can be aligned with high confidence, the rate is slightly lower (0.45 × 10−9 bp−1 year−1) and the level of divergence is 1.2% (13), implying an average time to the most common ancestor of 13 million years, assuming uniformity of the mutation rate over this time (95% ETPI 11 to 17 million years; table S11).
13 million years of the hominid line
This is not at all the first study to highlight the extreme dubiousness of the usual scholastic assumptions regarding the Pan-Homo divergence, which taint so many genetic studies, turning their chronological estimates totally worthless.
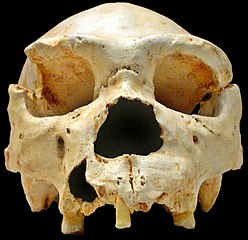
In 2010, Wilkinson et al. estimated a Pan-Homo divergence rate of 8-10 Ma. In 2012 Langergraber et al. recalibrated previous studies getting a Pan-Homo divergence bracket of 6.78-13.45 Ma (fig.2), while the divergence from Gorilla would be significantly older: 8.31-20.0.
 |
| Fig. 1 from Langergraber 2012. Legend: Diagram illustrating the branching pattern and timing of the splits between humans, chimpanzees, bonobos, western gorillas, and eastern gorillas. The paler shading indicates the range of split times inferred in this study. Cartoon skulls indicate approximate age of the indicated fossil remains, but do not imply that these fossils were necessarily on those ancestral lineages or that entire crania actually exist for these forms. |
A key fossil affecting this controversy is Sahelanthropus tchadiensis (Toumaï), which has been recently confirmed to be in the human line on several hardly questionable traits and is dated to c. 7 Ma.
A related debate is whether primates in general are much older than usually claimed and lived already in the Jurassic, something suggested by the already mentioned Wilkinson study and also by Heads 2010. Here a major issue is that mainline conservative estimates would have the ancestors of New World monkeys swimming (island hoping) to South America, something that those monkeys (and most other primates) simply will not do. The radiation of primates to South America and possibly also Madagascar is much better explained if these animals could just tree-hop, rather than island-hop to their destinations. However this would demand a radical revision of the usual age estimate for vertebrate radiation, what so far lacks fossil support (but lack of evidence is not evidence of lack, you know: fossil ages can only be taken as terminus ante quem dates and not absolute direct references).
But this is a side question, what really matters to us is that our ancestors split from the chimpanzee line c. 13 Ma (according to this study) and not after 8 Ma in any case (weighting all the evidence). This not just renders most "molecular clock" estimates useless and effectively false (wrong, erroneous, inadequate, misleading, junk, pseudoscientific...) but also help us to rethink our ancestral history in the African savannas since long before we became humans (Homo sp.)
Looking for some ecological context clues, I found this 1996 study by Jean Maley, which shows that Africa was largely humid in the early Miocene (smectite: evidence of water) but that it became increasingly arid towards the middle Miocene (kaolinite: evidence of sand). Up to this key ecological change of the Middle Miocene, the rainforest extended all the way to Egypt and East Africa. This kind of ecology allows for the common ancestor of African great apes to have arrived and first diverged in a jungle-dominated ecology and, later, for the speciation event leading to humans (bipedalism) to have happened as this once widespread jungle became scarcer, yielding to deserts and savanna.
 |
| Sahelanthropus (from fossilized.org) |
It just makes all sense that the evolution of bipedalism was coincident with the vanishing of that originally widespread jungle environment whose dating is of approx. 13 Ma ago. However it must be said that the consolidation of the Sahara only happened much later, c. 7 Ma ago, already approaching the Pliocene.
Regardless of the exact split-time, a big question I have on hominid evolution is how on Earth did our small-brained and small-toothed precursors like Toumaï survive in the open savannas and grasslands without fire nor weapons. Even if they resorted to trees (isolated or in patches) for refuge, there were already felines of the saber-toothed family roaming in Africa and these big cats were no doubt be able to climb on trees and in some cases they have been shown to predate on australopithecines. How could our precursors in the hominin line be able to face this menace without the advantage of speed (as ruminants have) or good defenses? Were their strong forelimbs, together with team action enough to confront the threat of predators? Did they use primitive weapons such as branches and stone throwing?