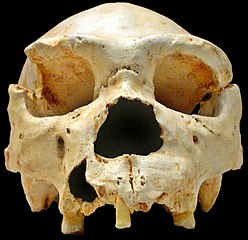
 |
| H. heidelbergensis from Atapuerca Cranium 5 "Miguelón" (CC by José Manuel Benito) |
About half an hour ago, somewhat cryptic comments in this blog and my email woke me up, more abruptly than I would have desired maybe, to a new game-breaking finding: researchers have sequenced the mtDNA of a 400,000 years old Homo heidelbergensis from Atapuerca (Iberian Peninsula, Europe) and it was not at all like most would have expected.
Mathhias Mayer et al., A mitochondrial genome sequence of a hominin from Sima de los Huesos. Nature 2013. Pay per view → LINK [doi:10.1038/nature12788]
Abstract
Excavations of a complex of caves in the Sierra de Atapuerca in northern Spain have unearthed hominin fossils that range in age from the early Pleistocene to the Holocene1. One of these sites, the ‘Sima de los Huesos’ (‘pit of bones’), has yielded the world’s largest assemblage of Middle Pleistocene hominin fossils2, 3, consisting of at least 28 individuals4 dated to over 300,000 years ago5. The skeletal remains share a number of morphological features with fossils classified as Homo heidelbergensis and also display distinct Neanderthal-derived traits6, 7, 8. Here we determine an almost complete mitochondrial genome sequence of a hominin from Sima de los Huesos and show that it is closely related to the lineage leading to mitochondrial genomes of Denisovans9, 10, an eastern Eurasian sister group to Neanderthals. Our results pave the way for DNA research on hominins from the Middle Pleistocene.
The key figure is this one, which phylogenetically relates the newly sequenced mtDNA with the known Homo ones:
 |
Figure 4: Bayesian phylogenetic tree of hominin mitochondrial relationships based on the Sima de los Huesos mtDNA sequence determined using the inclusive filtering criteria.
All nodes connecting the denoted hominin groups are supported with posterior probability of 1. The tree was rooted using chimpanzee and bonobo mtDNA genomes. The scale bar denotes substitutions per site. |
It has been argued by all sides (myself included) that the H. heidelbergensis of Atapuerca and other European locations are ancestral to Neanderthals. Some say that also to H. sapiens, while others argue that ours is a wholly distinct line, derived from H. rhodesiensis, and yet others claim that H. rhodesiensis is not different from H. heidelbergensis in spite of being older and rooted, it seems, in South Africa.
The clear evidence for migrations out of Africa, before our species, is limited to two periods: (1) the c. 1.8 Ma old migration of H. erectus/georgicus with Olduwayan technology (mode 1, "choppers"), and (2) the c. 1 Ma old migration of H. ergaster/antecessor (sometimes also confusingly called H. erectus) with Acheulean technology (mode 2, typically "hand axes"). Archaeological evidence for later migrations does not exist.
See: Late human evolution maps at Leherensuge.
So we could well ask, if H. heidelbergensis is not ancestral to Neanderthals, then where do Neanderthals come from?
It must be answered that we do not know yet if H. heidelbergensis is or not ancestral to Neanderthals or in what degree it is. The mitochodrial (maternal) lineage may well be misleading in this sense. Denisovans themselves were much more related to Neanderthals via autosomal (nuclear) DNA than the mtDNA, so it may also be the case with European Heidelbergensis.
In fact it is still possible that these individuals represent some sort of admixture between older and newer layers of human expansion. But there is no clear answer yet. What is clear is that no Neanderthals have these mitochondrial sequences but others closer to those of H. sapiens - and this is the most puzzling part in fact.
But one thing is clear: the World is much bigger than just Europe, and that was also the case back in Paleolithic times. Our answer may well lay under the sands of some tropical desert, the waters of the sea or whatever other place in Asia or Africa.
Even if we'd find the "missing link", so to say, we might not be able to discern it as such without genetic sequencing and that is often not even possible at all. However this pioneer research, as well as its precursors on a bear also from Atapuerca and a 700,000 years old horse (the true record of ancient DNA recovery), give us some hope of getting an improved, even if sometimes perplexing, understanding of the complexity of the human adventure.
Re the horse: The horse sample was frozen for most if not all of its geological time enabling excellent DNA conservation. As Atapuerca is in the temperate zone, the recovery of even mtDNA is an amazing surprise. Atapuerca is more representative of other early Homo sites, which does provoke great optimism. I had written off the tropical Flores specimens as hopeless for any DNA recovery, but maybe not?
ReplyDeleteFair enough, I guess.
DeleteSaid that it's plausible that the Atapuerca remains were also frozen for long: in the Ice Age the Northern Iberian Plateau (avg. height 700 m.) was quite inhospitable and if the remains were at the bottom of a natural pit ("sima") that would have also helped to keep the freezing conditions even when the surface lacked ice (pits were used in the past over here to keep ice well into summer: "neveros" or "neveras", from "nieve"=snow, a term still used for fridges). I don't know if it was the case but I would not be too surprised.
Anyhow there have been sequences achieved also in warm conditions like those of Australia (not so old but quite old anyhow), so I would not be too surprised if the old bones have still a lot of old stories to tell us. But maybe better in drier conditions than tropical rainforests, those seem the worst ones for preservation.
I can't say. I'm not really privy of the techniques and their limitations.
Somewhat off topic, but what do you think anyway...
ReplyDeletehttp://polishgenes.blogspot.com.au/2013/12/the-globular-amphora-man-from-late.html
What do you expect me to say? MtDNA K, lactose intolerant, longfaced... A single individual is not too informative.
DeleteYou say: "No doubt, his people were largely replaced by newcomers from the east and also west during the frequent population shifts in the region after the Neolithic"...
I wouldn't draw such conclusions on a single individual's data, not at all. In fact I'd expect in general Globular Amphorae people in general to be ancestral to modern populations of the area at least in some (or even large) degree. But you attempt to draw the opposite conclusions on a single data point, well... I cannot agree with such a risky undertaking.
At least you show some doubt because of the millet diet.
mtDNA K2a is actually more common in Poland than most other European countries today. But the millet is indeed the smoking gun for some serious population continuity in the region from the Chalcolithic to the present, because it was also enjoyed by the Wielbark people, and then right after them the earliest Polish Slavs.
Deletehttp://polishgenes.blogspot.com.au/2013/12/the-slavic-like-diet-of-wielbark.html
Nevertheless, I really can't see the present population of the North European Plain being largely derived of the Globular Amphora folk. Not after the Brandt et al. study.
I do not wish to entertain the off-topic issue for much longer but you are saying yourself that there are strong reasons for believing that "the present population of the North European Plain [I was mostly thinking of Poland specifically but whatever] being largely derived of the Globular Amphora folk".
DeleteNot sure which is the Brandt paper but there is a clear demographic peak in Poland upon the Kurgan (Baalberge, leading to Luboń→GA→CW) arrival. See: http://forwhattheywereweare.blogspot.com/2013/06/revisiting-demographics-of-northern-and.html
Instead in Denmark and Britain similar peaks correspond to Megalithism, while in Germany there is no such early peak but a depression and the later irregularities (the main "peak" or "plateau" being pre-CW) seem worth of a more detailed regionalized study.
Take this in account: consider the link above with some interest, please.
Maybe its time to strat to guess mtDNA of Red Deer Cave Man. My choice is between neanderthan and moder human.
ReplyDeleteIn fact it is still possible that these individuals represent some sort of admixture between older and newer layers of human expansion.
ReplyDeleteThat is what I think. While you are right that there is no documented ooA between ~1 million ya and ~100kya, we know that every 50-100ky climate is favorable for a bi-directional exchange. Moreover, during much of heidelbergensis (~800 - 300 kya from the continental science viewpoint), the climate was exceedingly mild in Europe, only getting colder after 400 kya.
Also, to me it very much looks like that there is the same development of cranial features and volume in Europe and Africa during that time frame, which indicates gene flow. Finally, there are no Neanderthals in the Levant or W Asia around 120 - 70 kya, which I find to be even a generous window for the most recent ooA. On the flip side, several finds - unfortunately mostly tool assemblages - just north of the Nubian-associated finds in Arabia, in W Asia, C Asia, and Siberia support continued occupation, there, which in my view can only be "non-Neanderthalized" heidelbergensis.
So, from that perspective, the Sima de los Huesos and Denisovan mtDNA could be more closely related to the European "erectus" (that from 1 million ya ooA) turned Eurasian heidelbergensis, while the Neanderthal/ sapiens lineage might be from the original African (ergaster) lineage, indicating subsequent gene flow into some heidelbergensis (that then became Neanderthal in Europe).
In the context of AMH admixture, this may mean that what is currently considered a Neanderthal component should really be heidelbergensis (they were only ~200 ky apart, 100 kya), and what is considered Denisovan admixture is really Asian erectus, if Denisova actually is heidelbergensis admixed with Asian erectus.
And the new find that E Asian populations also have "Denisovan" admixture makes this all-the-more likely. More importantly, E Asian introgression may explain a portion of the distinct extant E Asian phenotype, also since E Asian erectus would have had a long time to acquire cold adaptation.
Good to see you back posting, Maju. And yes, I realize you have posted about the possibility that "Denisovan" is really an Asian erectus signature, before.
... "we know that every 50-100ky climate is favorable for a bi-directional exchange."
DeleteDo we? Can we actually extrapolate the recent two pluvial and one semi-pluvial episodes to further into the past? I never read anything on that and I'm under the strong impression that not much is known about possible pluvial episodes before the Abbassia one (c. 125-90 Ka BP).
Whatever the case, any such migrations, especially if separated from the others by hundreds of millennia, should leave some sort of archaeologically noticeable track. Where is it?
"Also, to me it very much looks like that there is the same development of cranial features and volume in Europe and Africa during that time frame"...
There is a tendency to bigger brains (volume) but not really to similar cranial features. In fact our species H. sapiens has very distinctive cranial features compared to other Homo species, including Neanderthals: the skull is pulled upwards (higher vault, frontal bulge: http://forwhattheywereweare.blogspot.com/2013/01/frontal-bulge-almost-exclusive.html), the face flattened (much less prognathism), a chin appears, etc. Of course these features only show up since the first H. sapiens (Omo I, etc.), so it's debatable what happened earlier, but in any case I would not dare to say that our skull evolution is similar to that of other Homo species in the feature aspect, only in the volume one.
"... the Sima de los Huesos and Denisovan mtDNA could be more closely related to the European "erectus" (that from 1 million ya ooA) turned Eurasian heidelbergensis, while the Neanderthal/ sapiens lineage might be from the original African (ergaster) lineage, indicating subsequent gene flow into some heidelbergensis (that then became Neanderthal in Europe)".
But European Heidelbergensis has mode 2 (Acheulean), what should imply a gene flow from Africa, which is dated to maybe 900 Ka BP in the particular case of Iberia, either from Asia or directly from North Africa.
IF we follow only the mtDNA (forget Denisova by the moment because it may well have strong Neanderthal admixture, blurring the picture a lot when considerin nDNA), then the simplest explanation is that both are Heidelbergensis populations, somehow NOT leading to Neanderthals. The fact that there is no Neanderthal with such mtDNA lineages seems to underline that Heidelbergensis and Neanderthals are not related, at least not via mtDNA.
There is then the possibility of Neanderthals being a mixture of "pre-Sapiens" (H. rhodesiensis or whatever) and local Heidelbergensis but the data we have does not tell us about that at all.
One of the most intriguing bits is the Hathnora hominin, whose partial skull and humerus look very much Neanderthal (cf. Anek R. Sankhyan 2012: http://dx.doi.org/10.4236/aa.2012.23015), but is associated to Acheulean (mode 2) technologies. There is such a lack of palaeontological data from South Asia that it may really hold some key clues "in secret".
...
...
Delete"In the context of AMH admixture, this may mean that what is currently considered a Neanderthal component should really be heidelbergensis"...
It cannot be because the comparison was made directly with Neanderthals and IF Neanderthals are NOT derived from Heidelbergensis, then only the non-Neanderthal part of Denisova actually would tell us of Heidelbergensis admixture (limited to Native Australasians and such).
"... the new find that E Asian populations also have "Denisovan" admixture"...
Uh? That's new to me. All I've read only suggests a very thin pocket of "Denisovan" admixture in SE China, but so flimsy that is dubious and at best residual.
"E Asian introgression may explain a portion of the distinct extant E Asian phenotype, also since E Asian erectus would have had a long time to acquire cold adaptation."
I have no reason to believe that the East Asian phenotype has anything to do with "cold adaption", much less as the source of the population is very clearly in the South of the subcontinental region (tropical and subtropical climate).
I any case I have difficulty following you in all this. What you say could only make some sense assuming that Acheulean was actually spread by proto-Neanderthal (H. ergaster) populations but that it was partly spread by (mostly) cultural diffusion, maybe in admixed populations (pre-Neanderthals + H. heidelbergensis). Is that what you are saying or is it just in my mind?
DeleteMaju,
We have had this discussion of heidelbergensis and paleoclimate before.
The orbital forcing of the North African monsoon and corresponding E Mediterranean humidity has been well-established for the past several million years.
See http://en.wikipedia.org/wiki/North_African_climate_cycles and e.g. references 7, 8 and 12 therein, also https://www.era.lib.ed.ac.uk/bitstream/1842/560/2/CHAP_14.PDF. Over the past ~150,000 years, the correlation of proxy data, orbital forcing, and direct measures of increased humidity is so strong that I trust just the proxy data and orbital forcing for the period before then.
I don't know why you bring modern humans into the discussion: I was comparing European heidelbergensis with African specimen, which by some are also called heidelbergensis because of their similarity. From the metric studies I have seen, European heidelbergensis clusters with some African specimen both just based on the mandible, and also on overall cranial measures (and also clusters with a couple of Chinese skulls from 200 - 300kya: Dali and Jinniu Shan).
There are two lineages separated by something like a million years. One is represented by Sima de los Huesos and Denisova (over a ~300,000 y time frame), and the other by modern humans and all (but all very recent) Neanderthals studied, so far.
There is no explanation other than that Neanderthal has a more recent African lineage, (unless you want to suggest that extant Africans derive from a very early Eurasian heidelbergensis that also managed to survive in W Eurasia for a million years). In either case it shows more recent gene flow (by about a factor of two) than the populating of Europe by ergaster ~ 1 million ya, from which most people think European heidelbergensis derived.
(A different option is that the second lineage survived parallel in W Eurasia for about 1 million years, or survived yet further east and migrated west and eventually dominated Neanderthal. I find that extremely unlikely. A further option is that the relative mtDNA time clock is off by a factor of two, and the first split indicates the ~1.8 million years old erectus "georgicus" lineage, for which there is no further archeological record in Europe. Again, extremely unlikely.)
I don't think that an mtDNA lineage alone can question more than half a century of work that has shown it too be most likely that Neanderthal derived from heidelbergensis locally in western and central Europe. Neanderthal features are fewer and later in E Europe, and don't show up in the fossil record in the Levant or W Asia until ~60,000 ya, AFAIK.
The extreme cold after ~400kya, with just a short warm interval ~320kya, and an even shorter blip ~240kya, means that the total population in W/C Europe was small and would have been vulnerable to gene flow from presumably larger populations in more moderate climates from SE Europe to W and SW Asia (e.g., we have heidelbergensis finds in the Levant, with an associated complex <350 to 200kya - a period that mostly was not particularly dry). IMO the most straightforward explanation is that at one point this gene flow included a more recent lineage from Africa, and a late heidelbergensis/ very early Neanderthal population recovered with input from the southeast. It also makes sense that the W-E cline in Neanderthal features (some of which are cold adaptations) was actively sustained by a strong population in the milder SE.
As far as mode 2 Acheulean, I don't see where I wrote anything contradicting you or current understanding. I put European "erectus" in quotation marks to indicate its true derivation from African ergaster.
See this brief report for "Denisovan" admixture in E Asians:
http://www.nature.com/news/mystery-humans-spiced-up-ancients-sex-lives-1.14196
I find it much more plausible that we are talking about an E Asian erectus component, here, rather than some actual "Denisovan" population that lived 10,000 miles apart and in 10 different climate zones.
First, my apologies for the issue of climatic cycles. I probably did not understood your earlier explanations on the matter and I'm right now struggling to understand what the Kroon paper actually says. It is in any case very interesting and I thank you for your extra explanation here.
DeleteSecond....
"There is no explanation other than that Neanderthal has a more recent African lineage"...
That's exactly what it implies. The question is more recent than what? You say that more recent than H. ergaster's Acheulean expansion, and the graph seems to support your assumption, placing both Denisovans and Atapuercans in a line that split from hours some 800 Ka ago (estimated - it may need some adjustment towards 1 Ma or at least 900 Ka). The Atapuerca-Denisova split looks like some 600-700 Ka old and, in turn, the Neanderthal-Sapiens split looks like 500 Ka old only.
I must say that this simplistic interpretation may be way off, partly because it is unclear what exactly they are measuring but let's assume by the moment it is correct.
My big question is: what techno-cultural track shows the proto-Neanderthal migration out of Africa. Also: out of Africa but to where? To India, where we find the Narmada hominin, which looks totally Neanderthal but with Acheulean (not Mousterian) tech?
Re. H. erectus "asiaticus", with due caution, I would adventure now that it is not impossible that the H. ergaster actually replaced them and that the highly encephalized (almost modern) late Homo erectus of East Asia are actually derived from H. ergaster, at least largely. That would allow them to be in Indonesia at the time of the H. sapiens arrival. The lack of Acheulean East of the Mobius Line may be attributed to reasons such as the use of bamboo tools and/or the alleged lack of good quality stone in that area.
There is one instance of Acheulean in SE Asia but was short-lived.
The Nature article says:
"Reich said at the meeting that as well as interbreeding with the ancestors of Oceanians, they also bred with Neanderthals and the ancestors of modern humans in China and other parts of East Asia. Most surprisingly, Reich said, the genomes indicate that Denisovans interbred with yet another extinct population of archaic humans that lived in Asia more than 30,000 years ago — one that is neither human nor Neanderthal".
I guess you get your conclusions from the sentence "the ancestors of modern humans in China and other parts of East Asia", but this one is very confuse and probably refers, if anything, to the extremely low Denisovan admixture detected by one study only (i.e. dubious) in SE China. This hint of admixture could not be replicated in any non-dubious way, for example Wall 2013.
Even if it would be real, it'd be restricted to some very specific populations of SE China and would not be enough to expect it to have any obvious or large phenotype influence (10% level admixed people look almost invariably like the 90% of their ancestry, so less than 1% admixture should be quite irrelevant, more so as we carry double chromosomes and therefore two alleles of each gen).
The interesting part of that article is the suggestion of Denisovans being admixed with a third population, which is speculated to be an unknown Asian relative of Neanderthals (Hathnora?) But the information provided is extremely confuse, we'd have to see the paper as such but it does not seem to be published yet.
One change we have in Europe at a time roughly following this apparent gene exchange is the beginning of the transitions to Mousterian (~330kya) and heidelbergensis to Neanderthal (transition period ~ 300 - 150kya). Note that you can find two Africans with ~200ky mtDNA separation, so a divergence 500kya could easily affect another place ~200k years later.
ReplyDeleteIt's too late for Acheulean in S Asia, but fits with the Chinese Dali and Jinniu Shan, ~300 - 200kya. Now, I always associated changes during roughly that time in E Asia with late heidelbergensis expansion - but it is of course possible that whoever affected Europe also went directly, but separately, into Asia. On the flip side, if we consider Europe and at least the Near East as a population continuum, then there may not be much of a difference (an interpretation I prefer, given both Neanderthal and modern distribution ranges and time scales).
As to the Reich talk, my interpretation is that he wanted to point out how much better the very recent Denisova and Neanderthal genome sequences are, and that for that reason, E Asian "Denisovan" and "other" admixtures are now clearly apparent. Again, I find it more likely that these components are actually closer to heidelbergensis and E Asian erectus, but we will see.
"... whoever affected Europe also went directly, but separately, into Asia"...
DeleteNot sure what to think. But it's possible that proto-Neanderthals may have pushed Heidelbergensis eastwards. These were still "more evolved" and "brainier" than the first H. erectus, so they may be the reason why the East Asian archaic humans appear "more evolved" towards the end of the Middle Paleolithic.
One thing is clear: the Australasian distinctive admixture is Denisovan, what should read (at least partly) Heidelbergensis (and not Neanderthal or proto-Neanderthal). Anyhow I still think that Denisovans were about 50% Neanderthal (because of their autosomal affinity and because they lived side by side with Neanderthals) and therefore the real Heidelbergensis admixture in Native Australasians is of c. 2.3% and not 4.7% (i.e. Neanderthal admixture is being read twice).
I've long had a paradigm of [E. Asian H. Erectus--w/ shovel incisors and bamboo-hafted flake tools] vs. [African and W. Eurasian H. Ergaster > Antecessor > Heidelbergensis with handaxes (but not always)]. I have read that the "Acheulo-Yabrudians" of Israel, ca. 400k-300k bp, exhibited shovel teeth like E. Asians, as subsequently did some Neanderthals (weren't there as many as 5 or so possible "subspecies" of Neanderthal?) and, I believe, so did some H. Helmei.
ReplyDeleteTherefore, I pictured in the Yabrudian a E. and W. Eurasian hybridization and lithic tech exchange, leading to Africa's H. Helmei Lupemban (en route to later Stillbay and Moustero-Aterian) and Eurasia's Mousterian. I wonder if some un-Neanderthalized Heidelbergs survived in India, whose later descendants made it to southern Siberia and Southeast Asia (hence the Melanesia admixture).
As for your assertion of a lack of evidence for OOA in late-Middle/early-Upper Paleolithic times, I wonder if either Aterian or E. Africa Lakes harpoon cultures led to a spread of coastal village culture to the Persian Gulf and India, eventually giving rise to the earliest microblade cultures? (Perhaps related...just how old are the oldest Howiesonsport sites anyway? I keep reading drastically different estimates.) But I also believe there were some Cro-Magnon in W. Asia who settled for Neanderthalesque Mousterian technology (imitation?) for practical reasons, especially if they did not have large populations densities to support a large bank of blades to be distributed. You see such Mousteroid technology continue far to the northeast in Siberia, and perhaps the "pre-lithic horizon" finds in America.
I must say that shovel incisors (sinodonty) are also found in Africa, where no genetic connection with East Asia exists. I've seen them at least among Nuer and Oromos, and there are probably more. On the other hand SE Asians or Native Americans do not generally have them (sundadonty), so it's not neither an East Asian general trait nor even an Eurasian+ (non-African) specific one: much like epicanthic fold, it seems a matter of localized founder effects here and there.
DeleteOn the other hand straight hair is 100% non-African and I would (very tentatively) speculate of this trait as Neanderthal inherited, although it can have some other "internal" origin, of course. Sadly this trait can't be detected in ancient bones, nor can be in genetics so far, because the genetic causes are still unknown.
"I wonder if either Aterian or E. Africa Lakes harpoon cultures led to a spread of coastal village culture to the Persian Gulf and India, eventually giving rise to the earliest microblade cultures?"
All this corresponds with H. sapiens chronology, rather late in fact for Africa. It cannot be related with Neanderthals therefore, who are much older.
... "there were some Cro-Magnon in W. Asia who settled for Neanderthalesque Mousterian technology"...
Yes but as far as we know only in Palestine (with a possible mini-backflow to Egypt). It does not seem too central anymore to the Eurasian expansion of H. sapiens, because we known now of other MSA (and even the occasional Aterian tool) routes of migration via Arabia.
I'm not sure how familiar you are with all this so I'd recommend you to browse the category "out of Africa" in this blog and stop in the most relevant entries, because I have discussed this in the past a lot:
→ http://forwhattheywereweare.blogspot.com/search/label/out%20of%20Africa
More importantly maybe:
→ http://forwhattheywereweare.blogspot.com/2013/09/homo-sapiens-was-in-china-102000-years.html
→ http://forwhattheywereweare.blogspot.com/2013/07/homo-sapiens-from-central-china-dated.html
→ http://forwhattheywereweare.blogspot.com/2013/07/middle-paleolithic-industries-of.html
→ http://forwhattheywereweare.blogspot.com.es/2012/08/ancient-homo-sapiens-from-laos-46-63000.html
→ http://forwhattheywereweare.blogspot.com/2012/01/denisovan-admixture-may-actually-be.html
→ http://forwhattheywereweare.blogspot.com/2011/06/various-options-for-migration-out-of.html
→ http://forwhattheywereweare.blogspot.com/2011/01/coastal-route-through-arabia-130000.html
→ http://forwhattheywereweare.blogspot.com/2010/10/east-asian-jaw-from-100000-years-ago-is.html
→ http://forwhattheywereweare.blogspot.com/2010/10/some-key-archaeological-papers-on.html
So-called "shovel-shaped incisors" are extremely common among indigenous Americans (more so than among populations of eastern Asia on average) and also among Neanderthals.
DeleteAlright, I stand corrected on that detail (because I think that you really may be right after all). However I wonder if this is that way all through America or is somehow regionalized, because my first comment was based on my visual impression from South American Natives and you may instead be thinking of North American ones (just a guess).
DeleteWhat is clear in any case is that neither it is an invariable trait among East Asians (and derived populations) at all and that it is not exclusive of these either (but also found among populations unrelated to them - thinking of Nuer rather than Swedes here).
I just typed out a comment, and now I don't see it. Guess I should've copied in case of the need to re-paste.
ReplyDeleteThere's comment pre-moderation because I still have some stubborn trollish "fans" who I want out of this space for good. Let them write and bore people elsewhere with their rants.
DeleteSo it's only normal that comments do not show up straight away, because I have set it up to approve all comments before publication (and will remain that way for a while, even if I would prefer some sort of individualized treatment - but Blogger does not include such option).
However if your comment does not show up in a long time, say 24 hrs., email me please (or repost): most probably your comment went through but for no valid reason I did not get the notification email (or maybe I did not see your particular comment). My email is available at my profile (remove the anti-spam protection "DELETETHIS").
Sorry for whatever inconvenience.
Oh ok, it did post just fine. Sorry for the false alarm there.
ReplyDeleteOh ok, there it is, posted just fine. Sorry about the false alarm.
ReplyDeleteWith regard to OOA, what I am really proposing is a very early Erectus OOA to E. Asia, and a later Antecessor/Heidelberg OOA, followed by a E.-W.-hybridized Yabrudian spawning of both 1) Neanderthals moving northward and 2) Helmei moving southward "Back Into Africa" (BIA), leading to Africa's spectrum of modernizing traits (Irhoud, Omo, Stillbay...), then a brief OOA (Qafzeh), then another BIA retreat (mtDNA L3 perhaps?) during post-Toba ice age, followed by a late middle-Paleo. Aterioid/proto-Aurignacoid OOA (splitting into mtDNA M coast-walkers and mtDNA N long-hunters) admixing with a scattering of Toba survivors who had not gone BIA (Red Deer? Denisovan-mixed early moderns? Mungo?).
ReplyDeleteBut I will check out those links to get updated on your OOA reasonings. Thanks!
I perused most of your links where you "have discussed this in the past a lot." Thanks! So the big picture I gather is that you have Modern Humans existing far to the East at very early dates (100kBP+), using Middle Stone Age industries, and an East-to-West expansion around the beginning of the Upper Paleolithic. So do you see the transition from Middle to Upper Paleolithic in Eurasia (perhaps evidenced by, e.g., Petraglia's finds in India?) as being completely independent of and unconnected with Upper Paleolithic advances in Africa such as Stillbay, Aterian, Katanda, and Howiesonspoort? And one more question--I admit my mind tends to seize up and form a mental block with regard to vastly early MHS in China (100k+) because I have a hard time imagining what mtDNA/y-chrom. lineages they would be. M and C respectively, perhaps? Do those lineages diverge from L3 and CT, respectively, anywhere near that early? I did notice you don't put much stock in the molecular clocks. It just seems mind-boggling, hard to contemplate...
ReplyDelete"So the big picture I gather is that you have Modern Humans existing far to the East at very early dates (100kBP+)"...
DeleteThat seems to be the case now (most of the evidence is extremely recent and includes not just tools but also bones).
"So do you see the transition from Middle to Upper Paleolithic in Eurasia (perhaps evidenced by, e.g., Petraglia's finds in India?) as being completely independent of and unconnected with Upper Paleolithic advances in Africa such as Stillbay, Aterian, Katanda, and Howiesonspoort?"
I'm not sure which are those "Upper Paleolithic" advances of Africa you mean. Aterian for example is pretty much "Mousterian-like" (although different), i.e. mode 3, as is in general all the MSA. They indeed have some impressive advances like the heat retouch or the Katanda harpoons (so Magdalenian-like but a lot earlier!) but what classically defines the technological UP (mode 4) is the creation of blades and bladelets with a non-Levallois (non-flake) technology that eventually became very popular all around.
I've had some recent discussion on the origins and, with still many uncertainties, I've been informed that Iran may well be at the origin of the innovation as per the evidence available today, with the always open possibility that has further oriental origins in South Asia, where the research is probably not as advanced yet.
"I admit my mind tends to seize up and form a mental block with regard to vastly early MHS in China (100k+) because I have a hard time imagining what mtDNA/y-chrom. lineages they would be. M and C respectively, perhaps?"
My opinion on the matter is that all modern East Asian lineages (save yDNA Q) were already there before the arrival of UP via Siberia and that there was no substantial genetic change with the arrival of mode 4, except for the expansion of yDNA Q (which should be the vector) in the NE and America. The expansion of mode 4 in East Asia seems largely caused by cultural diffusion, although we cannot exclude some migrations within this macro-geographical frame (maybe the expansion of O to Sundaland?)
One data point is clear: 40 Ka ago, 10-20 Ka before the arrival of mode 4, people around Beijing already had an mtDNA R-derived lineage (a now extinct or very rare B4'5 sublineage). This and other data support the notion of the fundamentals of the Eurasian expansion being consolidated (barring local changes later on) before the UP, with the exception of West Eurasia and the Far North, which correspond to that period (and in the case of N1, a later one). The UP must therefore be considered less "essential" and more of an epiphenomenon, with only some genetic (demic) relevance, which is:
1. The bulk of West Eurasian colonization (leading to Neanderthal extinction eventually)
2. The proto-Amerindian y-DNA Q branch carrying the concept to East Asia and leading the colonization of the Far North (and later America).
3. Some spillover to NE Africa, from Egypt to the Horn (and maybe even as far south as Tanzania), corresponding to African yDNA J1 and R1b-V88 and mtDNA M1 among others. This may be related to the genesis of the LSA. A secondary spillover to NW Africa from Europe seems independent from this one (Solutrean period: mtDNA H, probably also V).
...
"I did notice you don't put much stock in the molecular clocks."
DeleteI strongly dislike them. In most cases they seem to have no empirical basis, probably because they have been victim of "pop-science" and scholasticism. For example (and this is just one aspect), while there are good grounds to think that the Pan-Homo split is at least 8 Ma old (8-13 Ma in Langergraber 2012 for instance), most "molecular clock" approaches insist on using shorter calibrations even as short as 5 Ma. These aprioristic assumptions totally pollute whatever validity the method may have. But even the methods themselves are largely arbitrary and full of problems (which are of different nature for yDNA and mtDNA).
I do not fully discard the possibility of a realistic molecular clock for yDNA (with correct archaeological and paleontological calibration always, one of the main systemic problems) but it must use full chromosome (or at least a large segment of it) and not STR microsatellite markers. This approach was performed some time ago (using PG1000 data) by another amateur geneticist but his problem was, IMO, in the calibration used (way too conservative). So I tried to correct such errors: http://forwhattheywereweare.blogspot.com/p/y-dna-ages.html
So far this is the best "MC" I can fathom, but many lineages seem underrepresented (leading to low end estimates). Also please scroll down at that page for the update because the first calibration I used was ~80Ka for the arrival to India and that must now be recalibrated to c. 100 Ka instead.
In such a dynamic scientific environment as are both population genetics and human prehistory (also advancing very fast, especially out of Europe) we can't afford to be scholastic: we must be extremely open minded (but also very critical) and ride the still moving wave and not fall behind it. This is not Newtonian mechanics, whose principles were established long ago and are essentially fixed: this is avant-guard science and what today is dogma for some tomorrow may be trash.
I generally pride to be able to spot the "trash" earlier than most and being vindicated as time passes - but of course I'm also subject to erring (who isn't?)
Well that is quite the paradigm shift to wrap my mind around.
ReplyDeleteIt makes me wonder if L3 (being so close to M and N) might represent a back-migration
from W. Asia (Qafzeh?). In other words, maybe a migration of L2'3'4'6 (with y-chr. B-T) out of Africa so SO EARLY that the L2(xL3-M-N) and B(xCT) have been lost by now.
"...all modern East Asian lineages (save yDNA Q) were already there before the arrival of UP via Siberia"
Why via Siberia, just curious?
I'm glad to help open your mind to new possibilities. For more detailed info about my opinion on the matter I'd suggest to search for "molecular clock" in this blog.
Delete"It makes me wonder if L3 (being so close to M and N) might represent a back-migration
from W. Asia"...
Nah. L3 has seven basal branches and five of them correspond exclusively (or almost) to Africa, so the origin should be in Africa, even if most probably not far from the Red Sea (I estimate that between Sudan and Ethiopia). These five branches also seem to be older in their coalescence than M and N, counting from the common root L3 (each "→" indicates a coding region transition):
→L3a (Ethiopia)
→L3b'f (mostly routed via Chad to West Africa)
→L3c'd (origin Sudan/Ethiopia, L3d routed via Chad to West Africa mostly)
→L3e'i'k'x (similar origin most likely, L3e to West Africa and Near East, L3i and L3x concentrated at or near Ethiopia, L3k apparently to NW Africa)
→→L3h (East and NE Africa)
→→→M (South Asia and eastwards, in essence)
→→→→→N (SE Asia maybe and then dynamic expansion in several directions, one of them Westwards)
The approx. distribution of the haplogroups taken (a bit hurriedly, I admit) from this old entry (based on Behar 2008, may be incomplete): http://leherensuge.blogspot.com/2010/03/reviewing-mtdna-l-lineages-notes-l3-l4.html
Mutations taken from the latest PhyloTree build: http://www.phylotree.org/tree/subtree_L3.htm
It would seem that L3 experienced a mild expansion (star-like structure with seven branches) at about the time of the early OoA migration into Arabia (c. 125 Ka ago) but before the Asian expansion as such began (c. 100 Ka ago).
"Why via Siberia, just curious?"
DeleteBecause a series of archaeological studies on that region strongly suggest that:
1. It was first populated by some archaic species ("Denisovans" first, Neanderthals later)
2. There was a replacement by (presumably) H. sapiens with Aurignacian-like technology at about the same time when we see the same happening in Europe (c. 47 Ka ago in Altai, since c. 49 Ka ago in Europe, some millennia earlier in West Asia - all them clearly part of the same wider process)
3. There was an expansion of the UP (stone blade, mode 4) technology to East Asia via the North (Mongolia, NW China and possibly other areas not yet documented in Southern Siberia) beginning c. 30 Ka ago.
4. Recently it has been confirmed that ~24 Ka BP (in a Gravettian-like context) people East of Altai (Western shore of Lake Baikal) were still genetically like those of West Eurasia or so it seems (Y-DNA R*, mtDNA U*). On the other side we still find locally dominant Y-DNA Q in areas near Mongolia in Neolithic times. Further West in Altai this West Eurasian affinity seems to persist until the Iron Age, when we see the first spread of Eastern mtDNA lineages in Central Asia. This affinity is not just genetic but also archaeological, cultural: we can say with great certainty that West Eurasia extended in most of Prehistory all the way to Altai (and occasionally even further East). Only at some uncertain point after the LGM, there was an expansion westwards from East Asia but restricted to the Taiga area whose main vector was yDNA N1.
Some bibliography and referenced older entries:
→ http://ejournal.anu.edu.au/index.php/bippa/article/viewFile/84/75 (Altai MP-UP transition)
→ https://journals.uair.arizona.edu/index.php/radiocarbon/article/view/4122/3547 (Paleolithic Siberia and the origin of Native Americans)
→ http://paleo.sscnet.ucla.edu/BrantCA2001.pdf (Mongolia and surroundings early UP)
→ http://forwhattheywereweare.blogspot.com/2013/12/the-malta-adna-findings.html (Mal'ta aDNA)
→ http://forwhattheywereweare.blogspot.com/2013/12/ancient-east-asian-y-dna-maps.html (China's Neolithic/Metals yDNA)
→ http://forwhattheywereweare.blogspot.com.es/2012/11/mitochondrial-snapshots-from-east-west.html (Central Asian Metal Ages' mtDNA)
→ http://forwhattheywereweare.blogspot.com.es/2013/07/a-review-of-haplogroup-n-y-dna.html (yDNA N review)
Oh yes, I agree with UP arriving via Siberia.
ReplyDeleteMy question "why Siberia" was based on what I now realize was my grammatical misunderstanding of your sentence "all modern East Asian lineages (save yDNA Q) were already there before the arrival of UP via Siberia".
I thought you were were saying the modern lineages were already there via Siberia. (before arrival of UP)... Got it now! It's the later arrival of UP that is via Siberia. Thanks.
Of course: I meant that the rest of lineages arrived via SE Asia within the wider context of the Asian (and Australasian) expansion of H. sapiens, long before the UP. I see no reason (and rather a lot of negative reasons) to imagine any H. sapiens W-E migrations via Siberia before the proto-Amerindian (UP) episode, signaled by yDNA Q1 (and less importantly some mtDNA X2).
Delete