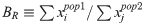This is really nice to read, considering that the archaeological data strongly favors a single out-of-Africa migration around that date (c. 125 Ka to Arabia and Palestine, see this, this and this among others, c. 100 Ka to South and East Asia) and that my own genetic modeling on mitochondrial DNA also fits that chronology (unlike most "molecular clock" scholastic rantings that are sold as "scientific truth" with no substantive backing whatsoever).
Admittedly the paper is not new (was published in February) but you know I have been missing important stuff with my information-overload stress crisis, so I'm making up now. Thanks to Ryan for bringing this up.
Martin Kuhlwilm et al., Ancient gene flow from early modern humans into Eastern Neanderthals. Nature 2016. Pay per view → LINK [doi:10.1038/nature16544]
AbstractIt has been shown that Neanderthals contributed genetically to modern humans outside Africa 47,000–65,000 years ago. Here we analyse the genomes of a Neanderthal and a Denisovan from the Altai Mountains in Siberia together with the sequences of chromosome 21 of two Neanderthals from Spain and Croatia. We find that a population that diverged early from other modern humans in Africa contributed genetically to the ancestors of Neanderthals from the Altai Mountains roughly 100,000 years ago. By contrast, we do not detect such a genetic contribution in the Denisovan or the two European Neanderthals. We conclude that in addition to later interbreeding events, the ancestors of Neanderthals from the Altai Mountains and early modern humans met and interbred, possibly in the Near East, many thousands of years earlier than previously thought.

As the title and the abstract say, Homo sapiens migrants out of Africa (i.e. into Asia and only later into its periphery) genetically influenced the branch of Neanderthals represented by the Altai specimen, what we can consider "Asian Neanderthals" but not the branch represented by El Sidrón and Vindija ("European Neanderthals"). This happened some 100 Ka ago, coincident with the archaeologically demonstrated dates for the out-of-Africa migration for our species and also likely Neanderthal-Sapiens hybrid fossils like Skhul-5 (right -- notice its lack of chin, a key and universal trait of H. sapiens, which allows us to give up with heavy browridges and other facial armature and still retain a strong bite, among other Neanderthaloid features, however it has a rounded and elevated skullcap with a high, almost vertical, forehead, a clear Sapiens trait).
The flow was in both directions. I do not have access to the paper itself but it is clear in the supplemental material (EDF-1). This hybridization event was distributed quite evenly among all "Greater Asians" or "non-Africans" in our species. The slightly lower score in French is surely caused by Neolithic admixture later on, bringing African-like genetics to Europe, which are absent in most of Asia, as well as in aboriginal Australasia and America.
Another apparent highlight in this paper (EDF-7) is that a second Neanderthal population, belonging to what I called above "European Neanderthals" (but related only to El Sidrón and not to Vindija) seems to have starred a second hybridization event affecting mostly Eastern populations (Han Chinese and Papuans in this paper's dataset). This, if confirmed, is quite unexpected and would require some explanation of the kind: there was a "European Neanderthal" population somewhere in Asia in the early times of Homo sapiens colonization and they got again admixed but this time affecting the derived populations in an irregular way. These irregularities would eventually be "flattened", I guess, at regional levels but for then the West Eurasian founders were out of the way.
A somewhat related recent paper, also mentioned by Ryan, is S. Sankararaman et al., The Combined Landscape of Denisovan and Neanderthal Ancestry in Present-Day Humans (Current Biology 2016), also pay-per-view, so judging on supplemental materials only. I must say I don't like this one that much but at least table S2 offers a summary of the state of the art of estimates of Neanderthal and "Denisovan" (H. heidelbergensis) admixture in a lot of populations (and not just three). It is apparent there that there is more Neanderthal admixture towards the East of Asia or "Greater Asia", what is very much counterintuitive and demands that a Neanderthal population (a "European Neanderthal" one per Kuhlwilm's data) existed somewhere towards the East of Asia, enabling for this secondary Neanderthal admixture event.
Perplexing maybe but that's what the data says. I wish we could find and sequence some of those Eastern Neanderthals which are so far just a genetic ghost, with the only possible known paleontological evidence being the Narmada skullcap, which is admittedly very much Neanderthal-like but is not associated in any way to Neanderthals' typical industry: the Mousterian, which has never been found east of Iran nor south of Mongolia.
Some have argued that the Zhirendong jaw, one of the key evidences for c. 100 Ka H. sapiens settlement of much of Asia, is a hybrid one, with clear H. sapiens traits (among others it has a chin) but also maybe "archaic" traits (among others its chin is rather small). If so, then we may be at the same time in this case before evidence of both the early migration of our species, Homo sapiens, to East Asia (or SE Asia, as it's quite to the south of China) and the second admixture event with Neanderthals, with those ghostly Oriental Neanderthals, related to El Sidrón ones in the Far West, quite paradoxically, that we have yet to properly identify.