Haplogroup N (Y-DNA) is spread from the Baltic to the South China Sea being one of those rare genetic links between East and West Eurasia (other than ultimate common ancestry) and one of the two Y-DNA lineages which expanded across the Northern Eurasian continent (the other one being Q).
While it is apparent to me and many others that the lineage originated in East Asia and expanded first Northwards to Siberia and later Westwards to Europe. I have found sometimes reluctance to accept this fact or difficulty understanding why. Some of the data of this paper may be of help in this regard. It is also a good exercise for those learning to understand how haploid genetics can be decoded into a meaningful pattern that reveals key parts of the untold history of peoples.
Hong Shi et al., Genetic Evidence of an East Asian Origin and Paleolithic Northward Migration of Y-chromosome Haplogroup N. PLoS ONE 2013. Open access → LINK [doi:10.1371/journal.pone.0066102]
Abstract
The Y-chromosome haplogroup N-M231 (Hg N) is distributed widely in eastern and central Asia, Siberia, as well as in eastern and northern Europe. Previous studies suggested a counterclockwise prehistoric migration of Hg N from eastern Asia to eastern and northern Europe. However, the root of this Y chromosome lineage and its detailed dispersal pattern across eastern Asia are still unclear. We analyzed haplogroup profiles and phylogeographic patterns of 1,570 Hg N individuals from 20,826 males in 359 populations across Eurasia. We first genotyped 6,371 males from 169 populations in China and Cambodia, and generated data of 360 Hg N individuals, and then combined published data on 1,210 Hg N individuals from Japanese, Southeast Asian, Siberian, European and Central Asian populations. The results showed that the sub-haplogroups of Hg N have a distinct geographical distribution. The highest Y-STR diversity of the ancestral Hg N sub-haplogroups was observed in the southern part of mainland East Asia, and further phylogeographic analyses supports an origin of Hg N in southern China. Combined with previous data, we propose that the early northward dispersal of Hg N started from southern China about 21 thousand years ago (kya), expanding into northern China 12–18 kya, and reaching further north to Siberia about 12–14 kya before a population expansion and westward migration into Central Asia and eastern/northern Europe around 8.0–10.0 kya. This northward migration of Hg N likewise coincides with retreating ice sheets after the Last Glacial Maximum (22–18 kya) in mainland East Asia.
Hong Shi has previously produced very interesting materials and this is no exception, however I find the use of chronological guesstimates as if these would be objective findings and treated as part of the central discourse (and not the mere side note where they belong) a bit nauseating and a cause of confusion.
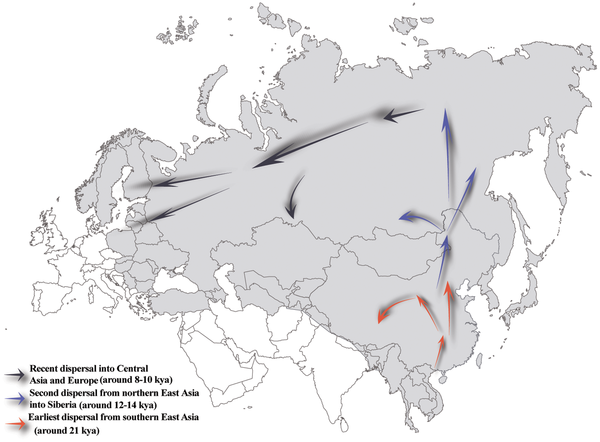 |
Figure 4. Proposed prehistoric migration routes for Hg N lineage.
(the pattern is correct but the dates are mere hunches, not any sort of objective facts)
|
Above we can see the reconstructed pattern of expansion of Y-DNA N in three phases.
In my understanding the dates are not way off, although I can only imagine that there is still room for improvement, especially regarding the "red" phase. After all NO may have split c. 60 Ka ago and the main branch, O, c. 50 Ka BP - and not the mere 25-30 Ka that Shi calculated (in a previous study but mentioned again here).
But the really interesting part is not molecular-clock-o-logy but this:
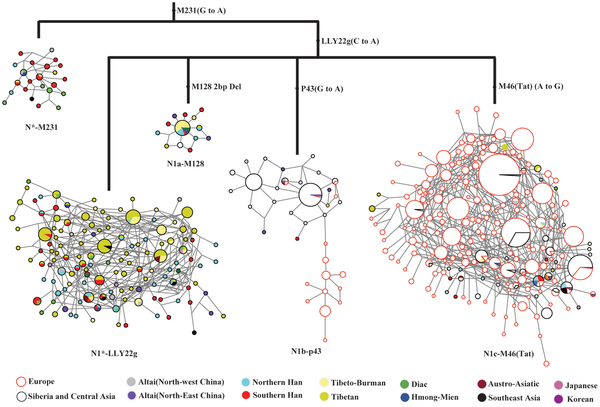 |
Figure 3. Median-joining networks for sub-haplogroups of Hg N lineage using Y-STR alleles.
The
diagnostic mutations used to classify the sub-haplogroups are labeled
on the tree branches. Each node represents a haplotype and its size is
proportional to the haplotype frequency, and the length of a branch is
proportional to the mutation steps. The colored areas indicate the
geographic origins of the studied populations or language groups. |
Here we can appreciate, with the labyrinthine limitations of the use of (too few?) STR markers, the apparent structure of the various haplogroups and paragroups under N. We can also see the STR diversity in numerical terms:
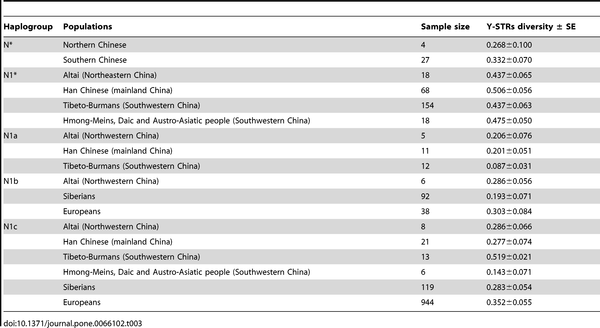 |
Table 3. Y-STRs diversity of Hg N sub-haplogroups.
|
Sadly the category "Han Chinese" is almost useless and one wonders why Shi et al. changed from the North/South polarity in the key paragroup N* to such a confusing terminology in N1.
In any case, it is quite evident that N arose in South China, spread, already as N1 to NE Asia and, later, some of that N1 (N1c mostly but also some N1b) spread Westwards reaching to Finland and other Eastern European populations. In the haplotype graph we can appreciate a distinct European-specific branch within N1b.
Update (Jul 28): some new findings (not considered in the study) and updated nomenclature.
See comments' section for greater details. Special thanks to Palamede for his efforts in clarifying the matter.
Commercial testing company FTDNA has recently detected some new markers within haplogroup N1 that alter the phylogeny. A synthesis of these findings can be seen in
this graph.
This new nomenclature was adopted by ISOGG but the study discussed here does not include it, using instead a 2011 nomenclature. Hence we must understand that:
- N* and N1* remain as such
- "N1a" (M128) is now known as N1c2a
- "N1b" (P43) is now N1c2b
- "N1c" (M46/Tat) is now N1c1
Therefore the N1 tree splits as:
- N1a (new clade, P189)
- N1b (new clade, L732)
- N1c (new clade including all previously named subhaplogroups)
- N1c1 (M46/Tat, former N1c)
- N1c2 (new clade, L666)
- N1c2a (M128, former N1a)
- N1c2b (P43, former N1b)
As far as I could gather, N1(xN1c) is so far only clearly represented by two FTDNA-tested singletons: a Slovakian (N1a) and someone of Polish surname (N1b1). However I may be missing some details. Whatever the case it is possible that, unless more samples show up in these groupings the tree may be later reverted to the original state (or something in between) because isolated individuals or families do not haplogroups make.
Also it is important to understand that commercial DNA testing companies have very unbalanced samples, clearly dominated by people of NW European (and to lesser extent other European) ancestry, what is not too useful when discerning what is where, producing sometimes the false impression of greater European diversity just because of greater number of samples.
On the other, hand the Hong Shi data reported above clearly shows a great number (and diversity) of East Asians within N1*, so the most likely conclusion is that the few Europeans within N1* are mere erratics within clades of East Asian origin, surely brought Westward by the overall N1 tide.
So in essence the conclusions of the paper remain unchallenged.















.jpg/320px-Rice_Plants_(IRRI).jpg)